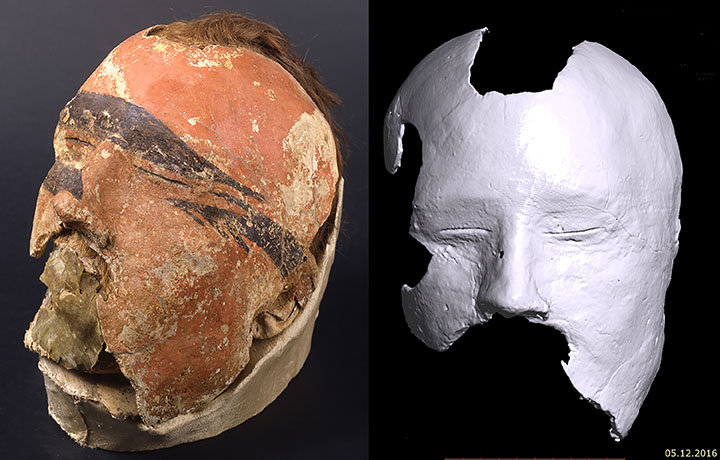
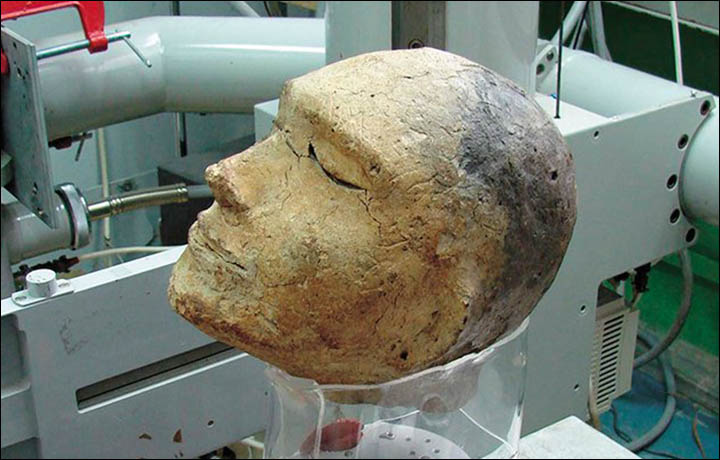
Rebecca Roberts introduces Gold of The Great Steppe, the first UK exhibition of recently excavated 2,700 year-old gold artefacts unearthed in remote burial mounds of East Kazakhstan. The Saka culture of Central Asia, flourishing from the 8th-3rd centuries BCE, was part of the Scythian world that dominated the Eurasian steppe zone from the Black Sea to Siberia. Located in the Altai mountain system, the Saka of East Kazakhstan were a vibrant society that occupied a landscape of open skies, rolling plains, winding rivers, creeping marshes, and soaring mountains. Based on discoveries made during an ongoing programme of excavation by archaeologists in the East Kazakhstan region, many of the artefacts were unearthed during the pandemic. They represent the resilience and determination of Kazakhstani archaeologists to protect and document their heritage, which is under threat from looting and degradation due to climate change. This exhibition is a partnership between the East Kazakhstan Regional Museum of Local History and the University of Cambridge. It will bring to the UK and display Kazakhstani archaeological finds and research from three different burial complexes in East Kazakhstan: Berel, Shilikti and Eleke Sazy. The show places archaeological finds discovered in the last three years by Kazakh archaeologists on a western global stage for the first time, amplifying voices that often go unheard in UK museums.
Sunday 31 October 2021
Friday 11 September 2020

Wednesday 9 September 2020
Islam, Literature and Society in Mongol Anatolia

Islam, Literature and Society in Mongol Anatolia

- ISBN-10 : 1108713483
- Paperback : 323 pages
- ISBN-13 : 978-1108713481
- Publisher : Cambridge University Press (12 Nov. 2020)
- Language: : English
From a Christian, Greek- and Armenian-speaking land to a predominantly Muslim and Turkish speaking one, the Islamisation of medieval Anatolia would lay the groundwork for the emergence of the Ottoman Empire as a world power and ultimately the modern Republic of Turkey. Bringing together previously unpublished sources in Arabic, Persian and Turkish, Peacock offers a new understanding of the crucial but neglected period in Anatolian history, that of Mongol domination, between c. 1240 and 1380. This represents a decisive phase in the process of Islamisation, with the popularisation of Sufism and the development of new forms of literature to spread Islam. This book integrates the study of Anatolia with that of the broader Islamic world, shedding new light on this crucial turning point in the history of the Middle East.
The Great Mongol Shahnameh
By Robert Hillenbrand

- Hardcover : 544 pages
- ISBN-10 : 1898113831
- ISBN-13 : 978-1898113836
- Product Dimensions : 30 x 15 x 32.7 cm
- Publisher : Yale University Press (23 Feb. 2021)
- Language: : English
A detailed study of the Great Mongol Shahnameh, considered to be the greatest of all Persian illustrated manuscripts The Great Mongol Shahnameh is widely considered to be the definitive version of Firdausi's epic poem, and the greatest of all Persian illustrated manuscripts. The paintings from this manuscript are held in private collections and institutions around the world, and have only been seen together in a single volume once since they were originally dispersed. This monograph reunites the paintings and reproduces them as 67 full-page, high quality color plates, alongside an analysis by leading scholar of Islamic art, Robert Hillenbrand. With newly commissioned photographs and insights into technical aspects of the paintings, The Great Mongol Shahnameh is a comprehensive resource for those interested in Persian art and manuscripts.
Tuesday 8 September 2020














Monday 15 June 2020
The Cultures of Ancient Xinjiang

The Cultures of Ancient Xinjiang, Western China: Crossroads of the Silk Roads
by
- Paperback: 218 pages
- Publisher: Archaeopress (19 Dec. 2019)
- Language: English
- ISBN-10: 178969406X
The Cultures of Ancient Xinjiang, Western China: Crossroads of the Silk Roads unveils the ancient secrets of Xinjiang, western China, one of the least known but culturally rich and complex regions located at the heart of Asia. Historically, Xinjiang has been the geographic hub of the Silk Roads, serving international links between cultures to the west, east, north and south. Trade, artefacts, foods, technologies, ideas, beliefs, animals and people have traversed the glacier covered mountain and desert boundaries. Perhaps best known for the Taklamakan desert, whose name translates in the Uyghur language as 'You can go in, you will never come out', here the region is portrayed as the centre of an ancient Bronze Age culture, revealed in the form of the famous Tarim Mummies and their grave goods. Three authoritative chapters by Chinese archaeologists appear here for the first time in English, giving international audiences direct access to the latest research ranging from the central-eastern Xiaohe region to the western valleys of the Bortala and Yili Rivers. Other contributions by European, Australian and Chinese archaeologists address the many complexities of the cultural exchanges that ranged from Mongolia, through to Kashgar, South Asia, Central Asia and finally Europe in pre-modern times.
Wednesday 6 May 2020
The Wonders of Creation and the Singularities of Painting

A Study of the Ilkhanid London Qazvini (Edinburgh Studies in Islamic Art)
by Stefano Carboni- Paperback: 456 pages
- Publisher: Edinburgh University Press (11 Feb. 2020)
- Language: English
- ISBN-10: 1474461395
Al-Qazwini's Wonders of Creation is one of a handful of extant illustrated codices produced under the Mongols of Persia. Al-Qazwini collected, edited and assembled a large body of literary works into a single text that reflects the cultural world of a medieval Arab encyclopaedist. In this lavishly illustrated volume, Stefan Carboni analyses the manuscript's miniatures, discusses the 368 paintings that illustrate the codex, and includes a partial critical translation of the related Arabic text. The codex contains a copy of a cosmographical text written in Arabic in Baghdad towards the end of the 13th century. The cosmography represents a physical description of the world arranged from the outer spheres of the universe, where the throne of God, the Angels and the Planets are located, down to Earth where the Peoples living in the Islands of the Oceans, the Mineral, the Vegetal and the Animal Kingdoms are described throughout the text. About the series: Edinburgh Studies in Islamic Art offers readers easy access to the most up-to-date research across the whole range of Islamic art, representing various parts of the Islamic world, media and approaches. Books in the series are beautifully illustrated academic monographs of intellectual distinction that mark a significant advance in the field.
Tuesday 5 May 2020
Uzbekistan: The Road to Samarkand

- by
- Hardcover: 336 pages
- Publisher: Assouline Publishing (February 26, 2020)
- Language: English
- ISBN-10: 1614288917
At the heart of Central Asia lies a land where colossal mountains and sweeping valleys sleep under a blanket of lush greenery. Crowned with golden palaces and wondrous monuments, the architectural landscape of the region is so rich with detail, the structures have been said to mirror the heavens themselves. One of the few destinations on Earth where imagination aligns with reality, Uzbekistan flourishes with unparalleled scenery and unforgotten traditions. The towns and cities are like ‘open museums’, each edifice offering a unique and intricate aesthetic, each a testament to diverse cultural influences and diverse periods of history. Nature and architecture have a unique relationship, seemingly inspired by each other, as if they were trying to to outdo each other with their beauty. Discover the beautiful colors, textures and flavors of this incredible culture and journey through the cities of the Silk Road and the lands of Alexander The Great with stunning original photography by Laziz Hamani.



Visualizing Dunhuang

Seeing, Studying, and Conserving the Caves
Publications of the Department of Art and Archaeology, Princeton University
Paperback
17 Nov. 2020
- Paperback: 400 pages
- Publisher: Princeton University Press (17 Nov. 2020)
- Language: English
- ISBN-10: 0691208166

A beautifully illustrated study of the caves at Dunhuang, exploring how this important Buddhist site has been visualized from its creation to today.
Situated at the crossroads of the northern and southern routes of the ancient silk routes in western China, Dunhuang is one of the richest Buddhist sites in the world, with more than 500 richly decorated cave temples constructed between the fourth and fourteenth centuries. The sculptures, murals, portable paintings, and manuscripts found in the Mogao and Yulin Caves at Dunhuang represent every aspect of Buddhism. From its earliest construction to the present, this location has been visualized by many individuals, from the architects, builders, and artists who built the caves to twentieth-century explorers, photographers, and conservators, as well as contemporary artists.
Visualizing Dunhuang: Seeing, Studying, and Conserving the Caves examines how the Lo Archive, a vast collection of photographs taken in the 1940s of the Mogao and Yulin Caves, inspires a broad range of scholarship. Lavishly illustrated with selected Lo Archive and modern photographs, the essays address three main areas
-Dunhuang as historical record, as site, and as art and art history.
Leading experts across three continents examine a wealth of topics, including expeditionary photography and cave architecture, to demonstrate the intellectual richness of Dunhuang. Diverse as they are in their subjects and methodologies, the essays represent only a fraction of what can be researched about Dunhuang. The high concentration of caves at Mogao and Yulin and their exceptional contents chronicle centuries of artistic styles, shifts in Buddhist doctrine, and patterns of political and private patronage-providing an endless source of material for future work.
Contributors include Neville Agnew, Dora Ching, Jun Hu, Annette Juliano, Richard Kent, Wei-Cheng Lin, Cary Liu, Maria Menshikova, Jerome Silbergeld, Roderick Whitfield, and Zhao Shengliang.
Published in association with the Tang Center for East Asian Art, Princeton University
Saturday 2 May 2020












{kind=link}
Monday 27 April 2020
The Cumans and Magyars

The History and Legacy of the Steppe Nomads Who Raided Europe Throughout the Late Middle Ages
- Paperback: 135 pages
- Publisher: Independently published (8 April 2020)
- Language: English
- ISBN-13: 979-8635405147
Before the Mongols rode across the steppes of Asia and Eastern Europe, the Cumans were a major military and cultural force that monarchs from China to Hungary and from Russia to the Byzantine Empire faced, often losing armies and cities in the process. The Cumans were a tribe of Turkic nomads who rode the steppes looking for plunder and riches, but they rarely stayed long after they got what they wanted.
From the late 9th century until the arrival of the Mongols in 1223, there was virtually nothing that could be done to stop the Cumans. Old Russian chronicles, Byzantine texts, Western European chronicles, and travel diaries of Islamic scholars all reveal that the Cumans were a threat to any kingdom in their path. Some kingdoms chose to fight the Cumans and often suffered heavy destruction, while others believed buying them off was the more reasonable course of action. The latter course often brought them into intimate contact with the most powerful kingdoms of medieval Eastern Europe before the Cumans were eventually replaced by the Mongols, with the remaining Cumans dispersing and integrating into various European and central Asian kingdoms in the 13th century. Many Cumans joined the Mongol Golden Horde and later became Muslims, while some helped found dynasties in Bulgaria, Hungary, and Romania.
The Cumans came from somewhat mysterious origins before they became the western vanguard of a massive nomadic horde that grew in ferocity and effectiveness as the centuries passed, but they were far more than mindless barbarians interested in violence alone. Although violence did play a major role in early Cuman culture, sources reveal they were also interested in diplomacy and eventually integrated with their sedentary neighbors. Archaeological discoveries further indicate that their culture was unique, complete with mythology and some art, but in the end, the Cumans disappeared as quickly as they appeared on the historical scene, much like other nomadic peoples before and after them.
Of all the steppe peoples in the medieval period, perhaps none were more important to European history than the Magyars. Like the Huns and Avars before them and the Cumans and Mongols after them, the Magyars burst into Europe as a destructive, unstoppable horde, taking whatever they wanted and leaving a steady stream of misery in their wake. They used much of the same tactics as the other steppe peoples and lived a similar, nomadic lifestyle. The Magyars also had many early cultural affinities with other steppe peoples, following a similar religion and ideas of kingship and nobility, among other things.
That said, as similar as the Magyars may have been to other steppe nomads before and after them, they were noticeably different in one way: the Magyars settled down and became a part of Europe and Western Civilization in the Middle Ages. The Magyars exploded onto the European cultural scene in the late 9th century as foreign marauders, but they made alliances with many important kingdoms in less than a century and established their own dynasty in the area, roughly equivalent to the modern nation-state of Hungary. After establishing themselves as a legitimate dynasty among their European peers, the Magyars formed a sort of cultural bridge between the Roman Catholic kingdoms of Western Europe and the Orthodox Christian kingdoms of Eastern Europe. Ultimately, the Magyars chose the Roman Catholic Church, thereby becoming a part of the West and tying their fate to it for the remainder of the Middle Ages.
The Cumans and Magyars: The History and Legacy of the Steppe Nomads Who Raided Europe Throughout the Late Middle Ages examines how the Cumans and Magyars became influential players in the region, and the influence they had.
Subscribe to:
Posts (Atom)